 Azərbaycanca
Azərbaycanca Беларускі
Беларускі Dansk
Dansk Deutsch
Deutsch Española
Española Français
Français Indonesia
Indonesia Italiana
Italiana 日本語
日本語 Қазақ
Қазақ Lietuvos
Lietuvos Nederlands
Nederlands Português
Português Русский
Русский සිංහල
සිංහල แบบไทย
แบบไทย Türkçe
Türkçe Українська
Українська 中國人
中國人 United State
United State Afrikaans
Afrikaans| Еукаріоти Час існування: Орозирій — наш час 1850–0 Ma | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Клітина еукаріотів | ||||||||||
| Біологічна класифікація | ||||||||||
| ||||||||||
Супергрупи та царства | ||||||||||
Посилання | ||||||||||
|
Еукаріо́ти (інколи евкаріо́ти, від грец. εύ- — повністю, добре й грец. κάρυον — ядро) або я́дерні (лат. Eukaryota Whittaker & Margulis, 1978) — домен одно- та багатоклітинних організмів, що характеризуються переважно полігеномними клітинами, морфологічно сформованим ядром та наявністю мембранних субклітинних органел.
Геноми еукаріотичної клітини представлені:
- а) ядерним геномом, зосередженим у ядрі і представленим ядерною ДНК;
- б) у більшості клітин мітохондріальним геномом, зосередженим у мітохондріях і представленим мітохондріальною ДНК;
- в) у деяких клітин пластидним (зокрема, хлоропластним) геномом, що розташовується у пластиді, і представлений хлоропластною ДНК (генофором);
- г) геномом нуклеоморфу, який виявлений лише у кількох відділах водоростей у надзвичайно цікавій ядроподібній структурі, розташованій між оболонками пластиди та особливою клітинною системою — хлоропластним ендоплазматичним ретикулумом. У нуклеоморфі виявлена власна, нуклеоморфна ДНК.
Двогеномні клітини, в яких є ядерний та мітохондріальний геноми, характерні для грибів та тварин; тригеномні — з ядерним, мітохондріальним та пластидним геномами — для майже всіх рослин; чотиригеномні — з ядерним, мітохондріальним, пластидним та нуклеоморфними геномами, виявлені у хлорарахніофітових та криптофітових водоростей.
Морфологія еукаріотичної клітини
Клітини прокаріотів (бактерій та архей) та еукаріотів на фенотипічному рівні схожі за наявністю ДНК та білок-синтезуючого апарату, представленого рибосомами; наявністю клітинних мембран; ферментних комплексів, що забезпечують процеси реплікації, транскрипції, трансляції та синтезу АТФ. До складу клітин прокаріотів та еукаріотів входять білки, жири, вуглеводи, нуклеїнові кислоти, мінеральні речовини та вода.
Відмінності в будові клітин прокаріотів та еукаріотів пов'язані, в першу чергу, з тим, що майже вся ДНК прокаріотів складається з однієї хромосоми, і, у випадку бактерій, не пов'язана з білками-гістонами (хоча навіть бактерії мають гістоноподібні білки). Еукаріоти, на відміну від прокаріотів, зазвичай не містять генів, організованих у великі оперони, процес цитокінезу (поділу ДНК) проходить за участю веретена поділу. Рибосоми у еукаріотів більші, ніж рибосоми прокаріотів (умовна вага прокаріотичних рибосом становить 70S, тоді як еукаріотичних — 80S). Багато клітин еукаріотів здатні до фаго- та піноцитозу, мають морфологічно оформлене ядро, часто також мітохондрії, пластиди, ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, пероксисоми, мають іншу структуру джгутиків і джгутикових моторів, мають клітинний центр з центріолями. Для багатьох клітин еукаріотів характерні мітоз, мейоз, повноцінний статевий процес, проте набагато нижча ймовірність горизонтального переносу генів. Зазвичай еукаріотичні клітини більші за розмірами, ніж прокаріотичні — їхній середній діаметр становить близько 5—20 мікрон, тоді як у бактерії та архей — 0,5—2 мікрона.
Гіпотези походження еукаріотів
За палеонтологічними даними, прокаріоти на нашій планеті виникли близько 3,2 млрд років тому, тоді як еукаріоти набагато молодші — їх вік становить лише близько 1,6 млрд років. Сьогодні в межах доменів бактерій та архей описано близько 40 тис. видів (хоча оцінки їх загальної кількості складають близько 10 млн видів), тоді як у домені еукаріотів — близько 1,7 млн видів.
Першим відомим евкаріотом є наземний організм Diskagma buttonii, гіпотетичний вид грибів, рештки якого було знайдено у Південній Африці.
Автогенетична гіпотеза
Згідно з цією гіпотезою, еукаріотична клітина утворилась з прокаріотичної внаслідок розвитку плазмалемою системи інвагінацій, які надалі замкнулись навколо ділянок плазмалеми з ферментами дихального ланцюга, фотосинтетичними пігментами, нуклеоїда, відповідно утворивши мітохондрії, пластиди та ядро. Розвиток системи інвагінацій мембран призвів також до виникнення ендоплазматичної сітки, комплексу Гольджі, лізосом та ін. Проте автогенетична гіпотеза не змогла пояснити чимало фактів. Наприклад, чому мітохондрії та хлоропласти містять власну ДНК і розмножуються поділом, чому в мітохондріях та хлоропластах наявні легкі прокаріотичні рибосоми, чому поровий апарат мітохондрій та пластид подібний до порового апарату прокаріот і відрізняється від порового апарату плазмалеми еукаріот, як виник мітоз та з'явились мікротрубочки та ін.
Ендосимбіотична гіпотеза
Наприкінці XIX ст. видатний ботанік та фізіолог рослин М. С. Цвєт, на основі спостережень поділу хлоропласту та ядерця у зеленої водорості Spirogyra висловив припущення, що клітини рослин, можливо, являють собою симбіотичний утвір із клітини-господаря та бактеріальних внутрішньоклітинних симбіонтів, які відомі як хлоропласти та ядерця. Це припущення у 20—40-х роках XX ст. було детально розроблено, в першу чергу, К. С. Мережковським у так звану ендосимбіотичну гіпотезу походження еукаріотичної клітини. Проте ідеї К. С. Мережковського не знайшли підтримки у широкого загалу біологів. Гіпотезу ендосимбіотичного походження еукаріотів називали «вражаючим витвором людської фантазії», «луна-парком», «непристойною темою у колі освічених біологів». У 60-х роках гіпотеза ендосимбіотичного походження еукаріотичної клітини була заново сформульована американською дослідницею Л. Маргеліс.
Згідно з гіпотезою Мережковського—Маргеліс, еукаріотична клітина виникла внаслідок кількох ендосимбіозів: гіпотетична прокаріотична анаеробна клітина, здатна до фагоцитозу, захопила, проте не перетравила, а зберегла в цитоплазмі, аеробну гетеротрофну бактерію, яка трансформувалась у мітохондрію. Далі клітина-господар, що містила мітохондрію, вступила в симбіоз із рухливою спірохетоподібною гетеротрофною бактерією, яка дала початок джгутику. Після цього внаслідок автогенетичного процесу, пов'язаного з утворенням глибоких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря утворилось ядро. Далі базальні тіла джгутиків трансформувались в центри організації веретена поділу, внаслідок чого виник спочатку мітоз, а потім його модифікація — мейоз, разом зі статевим процесом. Виникла перша гетеротрофна еукаріотична клітина. Нащадки цієї клітини, еволюція яких пішла по шляху вдосконалення фагоцитозу, дали початок тваринам. Ті ж нащадки, які еволюціонували в напрямку удосконалення осмотрофного живлення, дали початок грибам. Симбіоз гетеротрофної еукаріотичної клітини з ціанобактерією («синьо-зеленою водорістю») призвів до появи рослин, оскільки симбіотична ціанобактерія надалі трансформувалась у пластиду.
Синтетична гіпотеза


Численні перевірки ендосимбіотичної гіпотези, виконані з широким використанням електронної мікроскопії, мікрохімічних та молекулярно-генетичних методів, з одного боку, підтвердили гіпотезу ендосимбіотичного походження мітохондрій та пластид, з іншого — автогенетичну гіпотезу походження ядра та більшості одномембранних клітинних органел. Таким чином, на початку XXI ст. теорія походження еукаріотів сформувалась на основі синтезу обох гіпотез — як автогенетичної, так і ендосимбіотичної, і отримала назву синтетичної гіпотези походження еукаріотів.
Сьогодні вважають, що першим кроком на шляху виникнення еукаріотів було утворення у гіпотетичного прокаріотичного пращура численних внутрішніх інвагінацій плазмалеми, які, з одного боку, замкнули прокаріотичний нуклеоїд у двомембранну оболонку (тобто утворили морфологічно оформлене ядро), а з іншого — призвели до утворення ендоплазматичної сітки та похідного від неї комплексу Гольджі, а також травних вакуолей та їх похідних — лізосом.
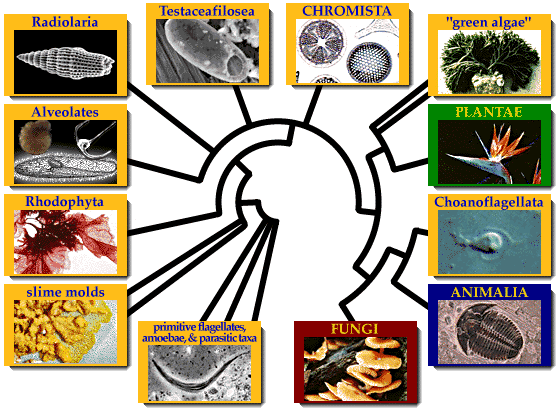
На другому етапі, внаслідок горизонтального переносу гену, що кодує білок тубулін від спірохетоподібних бактерій, примітивні еукаріоти набули здатності до синтезу тубулінових мікротрубочок. У результаті в еукаріотів з'явився цитоскелет, джгутики з базальним тілами, веретено поділу, мітоз. Базальні тіла джгутиків надалі у частини представників трансформувалися у клітинний центр, а порушення нормального мітозу (зокрема, скорочення інтерфази) призвели до виникнення мейозу і пов'язаного з ним статевого процесу. Група первинно безмітохондріальних еукаріотів сьогодні розглядається як перше, найпримітивніше царство еукаріотів — (, або ). Всі гіпохондріати є одноклітинними гетеротрофами з тваринною стратегією живлення.
На третьому етапі еукаріотична клітина утворила симбіотичний комплекс із прокаріотичною клітиною, схожою із сучасними альфа-протеобактеріями. Ця прокаріотична клітина надалі трансформувалась у мітохондрію. За результатами як цитологічних, так і молекулярних досліджень, найдавнішими мітохондріальними еукаріотами вважаються ті, які мають мітохондрії з особливими дископодібними кристами. Група мітохондріальних еукаріотів з дископодібними кристами прийнята як таксон рангу царства — Discicristata (Дискокристати). В основі царства дискокристат перебувають первинно гетеротрофні організми. На четвертому етапі еукаріоти розділилися на дві великі групи. Одна з цих груп має мітохондрії з трубчастими кристами, і складає царство (, трубчастокристні), друга — мітохондрії з переважно пластинчастими кристами, і приймається як царство Platycrsitates (платикристати, платівчастокристні). Подібно до дискокристат, в основі філ тубуло- та платикристат знаходяться первинно гетеротрофні організми із тваринною стратегією живлення.
На п'ятому етапі в еукаріотичному світі з'явилися перші рослини. За даними молекулярної та клітинної біології, ця подія пов'язана з симбіозом гетеротрофного еукаріота-платикристати з фотоавтотрофним прокаріотом — ціанобактерією (синьо-зеленою водорістю). Як наслідок цього симбіозу утворилась пластида, оточена двома мембранами, яка отримала назву первинно симбіотичної пластиди. Подальша дивергенція організмів із первинно симбіотичними пластидами обумовила виникнення у межах філи платикристат групи фотоавтотрофних відділів, які склали підцарство Plantae — рослини. Дивергенція гетеротрофних платикристат за типами живлення зумовила виникнення з одного боку, відділів з осмотрофним живленням — підцарства Fungi (гриби), з іншого — таксонів з фаготрофним живленням, що склали підцарство Animalia — тварини.
Велика радіація тубулокристат відбувалась майже паралельно з платикристатами, проте була пов'язана не стільки з еволюцією за типами живлення, скільки з еволюцією клітинних покривів та джгутикового апарату. В основі тубулокристат залишились амебоїдні організми, які складають підцарство Ameboflagellates (амебо-флогеляти). Дві прогресивніші філи представляють, з одного боку, таксони зі специфічними, альвеольованими покривами — Alveolata (альвеоляти), та з іншого — зі специфічними субмікроскопічними тричленними.
Первинні та вторинні ендосимбіози
Первинні ендосимбіози

Ендосимбіози, при яких в еукаріотичну клітину-хазяїна включався прокаріотичний симбіонт, що надалі трансформувався в клітинну органелу, називають первинними. На основі молекулярно-таксономічних досліджень наприкінці ХХ ст. було встановлено, що у світі еукаріотів первинні симбіози відбулися лише двічі. При першому ендосимбіозі прокаріотична бактерія, схожа з альфа-протеобактеріями, вступила в симбіоз із фаготрофним джгутиконосцем з царства гіпохондріат і трансформувалась у мітохондрію з трьома типами крист — дископодібними, трубчастими та пластинчастими. Подальша морфологічна еволюція мітохондрії відбувалася поступово, майже паралельно з клітиною-господарем. Так, у тубулокристат дископодібні кристи були редуковані, а збереглися трубчасті та інколи разом з ними — пластинчасті кристи. У платикристат були втрачені як дископодібні, так і трубчасті кристи, а збереглися лише пластинчасті (за винятком відділу Haptophyta).
Інший первинний ендосимбіоз відбувся між прокаріотичним оксигенним фотоавтотрофом — ціанобактерією (синьо-зеленою водорістю) та гетеротрофним організмом із царства платикристат. Як наслідок, виникла перша еукаріотична клітина, здатна до оксигенного фотосинтезу — так звана «протоводорість», тобто утворились рослини. У «протоводорості» пластида була вкрита двома мембранами, з яких внутрішня являла собою мембрану прокаріотичного ендосимбіонту — ціанобактерії (синьо-зеленої водорості), а зовнішня була похідною від плазмалеми клітини-господаря, і ймовірно являла собою видозмінену мембрану травної вакуолі, в яку потрапив ендосимбіонт.
«Протоводорість» дала початок трьом паралельним гілкам рослин із первинно симбіотичними пластидами — глаукоцистофітовим водоростям (Glaucocystophyta), червоним водоростям (Rhodophyta) та зеленим водоростям (Chlorophyta). Усі ці три відділи зберігають пластиди, що оточені лише двомембранною оболонкою. Глаукоцистофітові водорості зберігають цікаву атавістичну ознаку — між зовнішньою та внутрішньою мембраною пластиди розташовується шар муреїну — речовини, характерної для клітинних оболонок більшості евбактерій, і, зокрема, синьо-зелених водоростей. Відділ глаукоцистофітових є сліпою гілкою еволюції рослин[].
Пластиди червоних водоростей — родопласти — також зберігають деякі яскраві ознаки спорідненості із «синьо-зеленими водоростями» (ціанобактеріями), зокрема, так звані фікобілінові пігменти. Пластиди зелених водоростей — хлоропласти — зберігають найменше фенотипічних рис схожості із прокаріотичним ендосимбіонтом. Від зелених водоростей беруть початок наземні рослини, причому всі вони зберігають первинно симбіотичні хлоропласти.
Вторинні ендосимбіози
Червоні та зелені водорості неодноразово ставали ендосимбіонтами гетеротрофних еукаріотів із різних царств — дискокристат, тубулокристат та платикристат. Симбіози, що відбувалися за сценарієм «еукаріотичний хазяїн + еукаріотичний ендосимбіонт» називають вторинними. Внаслідок вторинних ендосимбіозів виникла велика група відділів водоростей із вторинно симбіотичними пластидами. Вторинно симбіотичні пластиди, на відміну від первинно симбіотичних, вкриті переважно чотирма мембранами, з яких внутрішня мембрана є залишком плазмалеми синьо-зеленої водорості (первинного ендосимбіонту), друга мембрана — видозміненою мембраною травної вакуолі господаря первинного ендосимбіонта (червоної або зеленої водорості), який є водночас вторинним ендосимбіонтом, третя мембрана — власне плазмалемою вторинного ендосимбіонта, четверта, зовнішня мембрана — плазмалемою господаря вторинного ендосимбіонта.
Цитоплазма, яка перебуває у проміжку між другою та третьою мембраною, так званий перипластидний простір, є редукованою цитоплазмою вторинного ендосимбіонта, і у деяких випадках зберігає 80S рибосоми та редуковану ендоплазматичну сітку. Зовнішні (третю та четверту) мембрани вторинно симбіотичних пластид називають хлоропластною ендоплазматичною сіткою.
Проте особливо цікавим є той факт, що у двох відділах водоростей із вторинно симбіотичними пластидами — хлорарахніофітових (Chlorarachniophyta) та криптофітових (Cryptophyta) — у перипластидному просторі зберігається навіть редуковане ядро вторинного ендосимбіонта — нуклеоморф. Нуклеоморф оточений двомембранною оболонкою і містить ДНК, яка (за наявними в нуклеоморфі генами) у хлорарахніофітових споріднена з ядерною ДНК зелених водоростей, а у криптофітових — із ядерною ДНК червоних водоростей.
У деяких водоростей, зокрема, в евгленових, вторинно симбіотичні пластиди вкриті не чотирма, а лише трьома мембранами. Вважається, що у цих водоростей одна з зовнішніх мембран пластиди була вторинно втрачена. Відомі випадки, коли пластиди (як первинно, так і вторинно симбіотичні) редукувалися, і тоді виникали вторинно гетеротрофні таксони. Зокрема, доведена вторинна редукція пластид у типу Apicomplexa, пращурами яких були фотоавтотрофні динофітові водорості (Dinophyta).
Є також дані, які дозволяють припустити, що оомікотові (Oomycota) утворились внаслідок вторинного переходу до гетеротрофного типу живлення від однієї з груп водоростей-тубулокристат. Крім того, вторинно гетеротрофні таксони низьких та середніх рангів (від видів до порядків і навіть класів) відомі майже в кожному відділі водоростей, а також серед вищих рослин[].
Поширення пластид через вторинні ендосимбіози обумовило виникнення значної кількості відділів водоростей. Наприклад, якщо водорості з первинно симбіотичними пластидами представлені лише трьома відділами (Glaucocystophyta, Rhodophyta, Chlorophyta), то водорості зі вторинно симбіотичними пластидами належать до 12 відділів. Серед них два відділи зберігають нуклеоморф (Chlorarachniophyta, Cryptophyta), 8 відділів мають чотиримембранні вторинно симбіотичні пластиди без нуклеоморфу, що походять від червоних водоростей (Raphidophyta, Chrysophyta, Eustigmatophyta, Xanthophyta, Phaeophyta, Bacillariophyta, Dictyochophyta, Haptophyta). Евгленофітові водорості мають тримембранні пластиди, пращурами яких є зелені водорості. У Dinophyta пластиди утворювались неодноразово, як внаслідок симбіозів із зеленими водоростями, так і навіть внаслідок третинних симбіозів із вторинно симбіотичними фотоавтотрофами.
Розмноження
Поділ клітини відбувається безстатево мітозом, процес дозволяє кожному дочірньому ядру отримувати по одній копії кожної хромосоми. Цикл життя багатьох еукаріотів включає статеву репродукцію, змінюючись з гаплоїдної фази, де в кожній клітині наявна одна копія кожної хромосоми, на диплоїдну фазу, де в кожній клітині наявні по дві копії кожної хромосоми. Диплоїдна фаза являє собою злиття двох гаплоїдних гамет, що формують зиготу, котра в свою чергу може ділитись в мітозі або пройти хромосомне скорочення в мейозі. У цьому шаблоні допустимі значні варіації — у тварин відсутня багатоклітинна гаплоїдна фаза, проте кожне покоління рослин може проходити через гаплоїдну та диплоїдну багатоклітинну фази.
В еукаріотів відношення площі до об'єму менше, ніж у прокаріотів, тому метаболічний процес та процес утворення клітини триває довше.
Еволюція процесу статевої репродукції — це первісна та фундаментальна характеристика еукаріотичних організмів. Базуючись на філогентичному аналізі, молекулярні біологи Дакс та Роджер припустили[], що статеве розмноження, хоч і не було основним способом розмноження, все ж було притаманне спільному предку всіх еукаріотів. Основний комплект генів, котрі фігурують на стадії мейозу, наявний навіть у таких паразитичних видів як Trichomonas vaginalis та Giardia intestinalis, котрі донедавна вважали асексуальними. Оскільки ці два види є нащадками родів, що на ранній стадії відділились від головного еволюційного дерева еукаріотів, було припущено, що основні гени, які фігурують в фазі мейозу, а отже і статеве розмноження, були, швидше за все, притаманними спільному предку еукаріотів. Еукаріотичні види, що помилково вважалися безстатевими, наприклад, паразит з роду Leishmania, все ж мають статевий цикл. Також, певні чинники вказують на те, що амеба, котра вважалась безстатевим видом, з давніх часів розмножувалася статевим шляхом, та що в більшості сучасних безстатевих видів нещодавно та незалежно один від одного прокинувся сексуальний потяг[][ ].
Джерела
- / відпов. редактори: Д. М. Гродзинський, . — Київ : КММ, 2012. — 744 с. — .
- Archibald, J.M., Cavalier-Smith, T., Maier, U. & Douglas, S. (2001). Molecular chaperones encoded by a reduced nucleus: the cryptomonad nucleomorph. J Mol Evol 52, 490501.
- Костіков І. Ю., Джаган В. В., Демченко Е. М., Бойко О. А., Бойко В. Р., Романенко П. О. Ботаніка. Водорості та гриби. — Київ, 2004.
- Thomas P. Curtis, William T. Sloan, and Jack W. Scannel (6). . PNAS. 99 (16): 10494—10499. Архів оригіналу за 24 вересня 2015. Процитовано 19 серпня 2008.
- Retallack, G.J., Krull, E.S., Thackray, G.D., and Parkinson, D. (2013). Problematic urn-shaped fossils from a Paleoproterozoic (2.2 Ga) paleosol in South Africa. Precambrian Research. 235: 71—87. doi:10.1016/j.precamres.2013.05.015.
- Сергей Ястребов (22.11.2017). . Новости науки на «Элементах». Архів оригіналу за 12 травня 2020. Процитовано 12 липня 2020.(рос.)
- P.J. Keeling, W.F. Doolittle Alpha-tubulin from early-diverging eukaryotic lineages and the evolution of the tubulin family. Molecular Biology and Evolution. 13: 1297—1305. doi:10.1093/oxfordjournals.molbev.a025576. PMID 8952074.
- Vossbrinck, C.R., J.V. Maddox, S. Friedman, B.A. Debrunner-Vossbrinck, and C.R. Woese (1987). Ribosomal RNA sequence suggests microsporidia are extremely ancient eukaryotes. Nature. 326: 411—414. doi:10.1038/326411a0. PMID 3550472..
Посилання
- http://mbe.oxfordjournals.org/cgi/reprint/13/10/1297
- http://itol.embl.de/itol.cgi [ 8 липня 2007 у Wayback Machine.]
- http://www.ucmp.berkeley.edu/alllife/eukaryota.gif [ 13 липня 2007 у Wayback Machine.]
- ЕУКАРІОТИ [ 12 березня 2016 у Wayback Machine.] // Фармацевтична енциклопедія
{kind=link}
 | Це незавершена стаття з біології. Ви можете проєкту, виправивши або дописавши її. |
Вікіпедія, Українська, Україна, книга, книги, бібліотека, стаття, читати, завантажити, безкоштовно, безкоштовно завантажити, mp3, відео, mp4, 3gp, jpg, jpeg, gif, png, малюнок, музика, пісня, фільм, книга, гра, ігри, мобільний, телефон, android, ios, apple, мобільний телефон, samsung, iphone, xiomi, xiaomi, redmi, honor, oppo, nokia, sonya, mi, ПК, web, Інтернет
Eukarioti Chas isnuvannya Orozirij nash chas 1850 0 Ma Fan Proterozoj Arhej Gadej Klitina eukariotiv Biologichna klasifikaciya Domen Eukarioti Eukaryota Whittaker amp Margulis 1978 Supergrupi ta carstva Archaeplastida Plantae Hacrobia SAR Stramenopiles Alveolata Rhizaria Excavata Amoebozoa Opisthokonta Animalia Fungi Posilannya Vikishovishe Eukaryota Vikividi Eukaryota EOL 2908256 NCBI 2759 Fossilworks 306691 Eukario ti inkoli evkario ti vid grec ey povnistyu dobre j grec karyon yadro abo ya derni lat Eukaryota Whittaker amp Margulis 1978 domen odno ta bagatoklitinnih organizmiv sho harakterizuyutsya perevazhno poligenomnimi klitinami morfologichno sformovanim yadrom ta nayavnistyu membrannih subklitinnih organel Genomi eukariotichnoyi klitini predstavleni a yadernim genomom zoseredzhenim u yadri i predstavlenim yadernoyu DNK b u bilshosti klitin mitohondrialnim genomom zoseredzhenim u mitohondriyah i predstavlenim mitohondrialnoyu DNK v u deyakih klitin plastidnim zokrema hloroplastnim genomom sho roztashovuyetsya u plastidi i predstavlenij hloroplastnoyu DNK genoforom g genomom nukleomorfu yakij viyavlenij lishe u kilkoh viddilah vodorostej u nadzvichajno cikavij yadropodibnij strukturi roztashovanij mizh obolonkami plastidi ta osoblivoyu klitinnoyu sistemoyu hloroplastnim endoplazmatichnim retikulumom U nukleomorfi viyavlena vlasna nukleomorfna DNK Dvogenomni klitini v yakih ye yadernij ta mitohondrialnij genomi harakterni dlya gribiv ta tvarin trigenomni z yadernim mitohondrialnim ta plastidnim genomami dlya majzhe vsih roslin chotirigenomni z yadernim mitohondrialnim plastidnim ta nukleomorfnimi genomami viyavleni u hlorarahniofitovih ta kriptofitovih vodorostej Morfologiya eukariotichnoyi klitiniKlitini prokariotiv bakterij ta arhej ta eukariotiv na fenotipichnomu rivni shozhi za nayavnistyu DNK ta bilok sintezuyuchogo aparatu predstavlenogo ribosomami nayavnistyu klitinnih membran fermentnih kompleksiv sho zabezpechuyut procesi replikaciyi transkripciyi translyaciyi ta sintezu ATF Do skladu klitin prokariotiv ta eukariotiv vhodyat bilki zhiri vuglevodi nukleyinovi kisloti mineralni rechovini ta voda Vidminnosti v budovi klitin prokariotiv ta eukariotiv pov yazani v pershu chergu z tim sho majzhe vsya DNK prokariotiv skladayetsya z odniyeyi hromosomi i u vipadku bakterij ne pov yazana z bilkami gistonami hocha navit bakteriyi mayut gistonopodibni bilki Eukarioti na vidminu vid prokariotiv zazvichaj ne mistyat geniv organizovanih u veliki operoni proces citokinezu podilu DNK prohodit za uchastyu veretena podilu Ribosomi u eukariotiv bilshi nizh ribosomi prokariotiv umovna vaga prokariotichnih ribosom stanovit 70S todi yak eukariotichnih 80S Bagato klitin eukariotiv zdatni do fago ta pinocitozu mayut morfologichno oformlene yadro chasto takozh mitohondriyi plastidi endoplazmatichnij retikulum kompleks Goldzhi lizosomi peroksisomi mayut inshu strukturu dzhgutikiv i dzhgutikovih motoriv mayut klitinnij centr z centriolyami Dlya bagatoh klitin eukariotiv harakterni mitoz mejoz povnocinnij statevij proces prote nabagato nizhcha jmovirnist gorizontalnogo perenosu geniv Zazvichaj eukariotichni klitini bilshi za rozmirami nizh prokariotichni yihnij serednij diametr stanovit blizko 5 20 mikron todi yak u bakteriyi ta arhej 0 5 2 mikrona Gipotezi pohodzhennya eukariotivZa paleontologichnimi danimi prokarioti na nashij planeti vinikli blizko 3 2 mlrd rokiv tomu todi yak eukarioti nabagato molodshi yih vik stanovit lishe blizko 1 6 mlrd rokiv Sogodni v mezhah domeniv bakterij ta arhej opisano blizko 40 tis vidiv hocha ocinki yih zagalnoyi kilkosti skladayut blizko 10 mln vidiv todi yak u domeni eukariotiv blizko 1 7 mln vidiv Pershim vidomim evkariotom ye nazemnij organizm Diskagma buttonii gipotetichnij vid gribiv reshtki yakogo bulo znajdeno u Pivdennij Africi Avtogenetichna gipoteza Zgidno z ciyeyu gipotezoyu eukariotichna klitina utvorilas z prokariotichnoyi vnaslidok rozvitku plazmalemoyu sistemi invaginacij yaki nadali zamknulis navkolo dilyanok plazmalemi z fermentami dihalnogo lancyuga fotosintetichnimi pigmentami nukleoyida vidpovidno utvorivshi mitohondriyi plastidi ta yadro Rozvitok sistemi invaginacij membran prizviv takozh do viniknennya endoplazmatichnoyi sitki kompleksu Goldzhi lizosom ta in Prote avtogenetichna gipoteza ne zmogla poyasniti chimalo faktiv Napriklad chomu mitohondriyi ta hloroplasti mistyat vlasnu DNK i rozmnozhuyutsya podilom chomu v mitohondriyah ta hloroplastah nayavni legki prokariotichni ribosomi chomu porovij aparat mitohondrij ta plastid podibnij do porovogo aparatu prokariot i vidriznyayetsya vid porovogo aparatu plazmalemi eukariot yak vinik mitoz ta z yavilis mikrotrubochki ta in Endosimbiotichna gipoteza Dokladnishe Endosimbiotichna teoriya Naprikinci XIX st vidatnij botanik ta fiziolog roslin M S Cvyet na osnovi sposterezhen podilu hloroplastu ta yadercya u zelenoyi vodorosti Spirogyra visloviv pripushennya sho klitini roslin mozhlivo yavlyayut soboyu simbiotichnij utvir iz klitini gospodarya ta bakterialnih vnutrishnoklitinnih simbiontiv yaki vidomi yak hloroplasti ta yadercya Ce pripushennya u 20 40 h rokah XX st bulo detalno rozrobleno v pershu chergu K S Merezhkovskim u tak zvanu endosimbiotichnu gipotezu pohodzhennya eukariotichnoyi klitini Prote ideyi K S Merezhkovskogo ne znajshli pidtrimki u shirokogo zagalu biologiv Gipotezu endosimbiotichnogo pohodzhennya eukariotiv nazivali vrazhayuchim vitvorom lyudskoyi fantaziyi luna parkom nepristojnoyu temoyu u koli osvichenih biologiv U 60 h rokah gipoteza endosimbiotichnogo pohodzhennya eukariotichnoyi klitini bula zanovo sformulovana amerikanskoyu doslidniceyu L Margelis Zgidno z gipotezoyu Merezhkovskogo Margelis eukariotichna klitina vinikla vnaslidok kilkoh endosimbioziv gipotetichna prokariotichna anaerobna klitina zdatna do fagocitozu zahopila prote ne peretravila a zberegla v citoplazmi aerobnu geterotrofnu bakteriyu yaka transformuvalas u mitohondriyu Dali klitina gospodar sho mistila mitohondriyu vstupila v simbioz iz ruhlivoyu spirohetopodibnoyu geterotrofnoyu bakteriyeyu yaka dala pochatok dzhgutiku Pislya cogo vnaslidok avtogenetichnogo procesu pov yazanogo z utvorennyam glibokih invaginacij plazmalemi navkolo nukleoyidu klitini gospodarya utvorilos yadro Dali bazalni tila dzhgutikiv transformuvalis v centri organizaciyi veretena podilu vnaslidok chogo vinik spochatku mitoz a potim jogo modifikaciya mejoz razom zi statevim procesom Vinikla persha geterotrofna eukariotichna klitina Nashadki ciyeyi klitini evolyuciya yakih pishla po shlyahu vdoskonalennya fagocitozu dali pochatok tvarinam Ti zh nashadki yaki evolyucionuvali v napryamku udoskonalennya osmotrofnogo zhivlennya dali pochatok gribam Simbioz geterotrofnoyi eukariotichnoyi klitini z cianobakteriyeyu sino zelenoyu vodoristyu prizviv do poyavi roslin oskilki simbiotichna cianobakteriya nadali transformuvalas u plastidu Sintetichna gipoteza Shema genetichnih zv yazkiv mizh carstvami organichnogo svitu Rozvitok zhivih organizmiv Chislenni perevirki endosimbiotichnoyi gipotezi vikonani z shirokim vikoristannyam elektronnoyi mikroskopiyi mikrohimichnih ta molekulyarno genetichnih metodiv z odnogo boku pidtverdili gipotezu endosimbiotichnogo pohodzhennya mitohondrij ta plastid z inshogo avtogenetichnu gipotezu pohodzhennya yadra ta bilshosti odnomembrannih klitinnih organel Takim chinom na pochatku XXI st teoriya pohodzhennya eukariotiv sformuvalas na osnovi sintezu oboh gipotez yak avtogenetichnoyi tak i endosimbiotichnoyi i otrimala nazvu sintetichnoyi gipotezi pohodzhennya eukariotiv Sogodni vvazhayut sho pershim krokom na shlyahu viniknennya eukariotiv bulo utvorennya u gipotetichnogo prokariotichnogo prashura chislennih vnutrishnih invaginacij plazmalemi yaki z odnogo boku zamknuli prokariotichnij nukleoyid u dvomembrannu obolonku tobto utvorili morfologichno oformlene yadro a z inshogo prizveli do utvorennya endoplazmatichnoyi sitki ta pohidnogo vid neyi kompleksu Goldzhi a takozh travnih vakuolej ta yih pohidnih lizosom Na drugomu etapi vnaslidok gorizontalnogo perenosu genu sho koduye bilok tubulin vid spirohetopodibnih bakterij primitivni eukarioti nabuli zdatnosti do sintezu tubulinovih mikrotrubochok U rezultati v eukariotiv z yavivsya citoskelet dzhgutiki z bazalnim tilami vereteno podilu mitoz Bazalni tila dzhgutikiv nadali u chastini predstavnikiv transformuvalisya u klitinnij centr a porushennya normalnogo mitozu zokrema skorochennya interfazi prizveli do viniknennya mejozu i pov yazanogo z nim statevogo procesu Grupa pervinno bezmitohondrialnih eukariotiv sogodni rozglyadayetsya yak pershe najprimitivnishe carstvo eukariotiv abo Vsi gipohondriati ye odnoklitinnimi geterotrofami z tvarinnoyu strategiyeyu zhivlennya Na tretomu etapi eukariotichna klitina utvorila simbiotichnij kompleks iz prokariotichnoyu klitinoyu shozhoyu iz suchasnimi alfa proteobakteriyami Cya prokariotichna klitina nadali transformuvalas u mitohondriyu Za rezultatami yak citologichnih tak i molekulyarnih doslidzhen najdavnishimi mitohondrialnimi eukariotami vvazhayutsya ti yaki mayut mitohondriyi z osoblivimi diskopodibnimi kristami Grupa mitohondrialnih eukariotiv z diskopodibnimi kristami prijnyata yak takson rangu carstva Discicristata Diskokristati V osnovi carstva diskokristat perebuvayut pervinno geterotrofni organizmi Na chetvertomu etapi eukarioti rozdililisya na dvi veliki grupi Odna z cih grup maye mitohondriyi z trubchastimi kristami i skladaye carstvo trubchastokristni druga mitohondriyi z perevazhno plastinchastimi kristami i prijmayetsya yak carstvo Platycrsitates platikristati plativchastokristni Podibno do diskokristat v osnovi fil tubulo ta platikristat znahodyatsya pervinno geterotrofni organizmi iz tvarinnoyu strategiyeyu zhivlennya Na p yatomu etapi v eukariotichnomu sviti z yavilisya pershi roslini Za danimi molekulyarnoyi ta klitinnoyi biologiyi cya podiya pov yazana z simbiozom geterotrofnogo eukariota platikristati z fotoavtotrofnim prokariotom cianobakteriyeyu sino zelenoyu vodoristyu Yak naslidok cogo simbiozu utvorilas plastida otochena dvoma membranami yaka otrimala nazvu pervinno simbiotichnoyi plastidi Podalsha divergenciya organizmiv iz pervinno simbiotichnimi plastidami obumovila viniknennya u mezhah fili platikristat grupi fotoavtotrofnih viddiliv yaki sklali pidcarstvo Plantae roslini Divergenciya geterotrofnih platikristat za tipami zhivlennya zumovila viniknennya z odnogo boku viddiliv z osmotrofnim zhivlennyam pidcarstva Fungi gribi z inshogo taksoniv z fagotrofnim zhivlennyam sho sklali pidcarstvo Animalia tvarini Velika radiaciya tubulokristat vidbuvalas majzhe paralelno z platikristatami prote bula pov yazana ne stilki z evolyuciyeyu za tipami zhivlennya skilki z evolyuciyeyu klitinnih pokriviv ta dzhgutikovogo aparatu V osnovi tubulokristat zalishilis ameboyidni organizmi yaki skladayut pidcarstvo Ameboflagellates amebo flogelyati Dvi progresivnishi fili predstavlyayut z odnogo boku taksoni zi specifichnimi alveolovanimi pokrivami Alveolata alveolyati ta z inshogo zi specifichnimi submikroskopichnimi trichlennimi Pervinni ta vtorinni endosimbioziPervinni endosimbiozi Shema evolyuciyi eukariotiv z pervinnimi ta vtorinnimi endosimbiozami Endosimbiozi pri yakih v eukariotichnu klitinu hazyayina vklyuchavsya prokariotichnij simbiont sho nadali transformuvavsya v klitinnu organelu nazivayut pervinnimi Na osnovi molekulyarno taksonomichnih doslidzhen naprikinci HH st bulo vstanovleno sho u sviti eukariotiv pervinni simbiozi vidbulisya lishe dvichi Pri pershomu endosimbiozi prokariotichna bakteriya shozha z alfa proteobakteriyami vstupila v simbioz iz fagotrofnim dzhgutikonoscem z carstva gipohondriat i transformuvalas u mitohondriyu z troma tipami krist diskopodibnimi trubchastimi ta plastinchastimi Podalsha morfologichna evolyuciya mitohondriyi vidbuvalasya postupovo majzhe paralelno z klitinoyu gospodarem Tak u tubulokristat diskopodibni kristi buli redukovani a zbereglisya trubchasti ta inkoli razom z nimi plastinchasti kristi U platikristat buli vtracheni yak diskopodibni tak i trubchasti kristi a zbereglisya lishe plastinchasti za vinyatkom viddilu Haptophyta Inshij pervinnij endosimbioz vidbuvsya mizh prokariotichnim oksigennim fotoavtotrofom cianobakteriyeyu sino zelenoyu vodoristyu ta geterotrofnim organizmom iz carstva platikristat Yak naslidok vinikla persha eukariotichna klitina zdatna do oksigennogo fotosintezu tak zvana protovodorist tobto utvorilis roslini U protovodorosti plastida bula vkrita dvoma membranami z yakih vnutrishnya yavlyala soboyu membranu prokariotichnogo endosimbiontu cianobakteriyi sino zelenoyi vodorosti a zovnishnya bula pohidnoyu vid plazmalemi klitini gospodarya i jmovirno yavlyala soboyu vidozminenu membranu travnoyi vakuoli v yaku potrapiv endosimbiont Protovodorist dala pochatok trom paralelnim gilkam roslin iz pervinno simbiotichnimi plastidami glaukocistofitovim vodorostyam Glaucocystophyta chervonim vodorostyam Rhodophyta ta zelenim vodorostyam Chlorophyta Usi ci tri viddili zberigayut plastidi sho otocheni lishe dvomembrannoyu obolonkoyu Glaukocistofitovi vodorosti zberigayut cikavu atavistichnu oznaku mizh zovnishnoyu ta vnutrishnoyu membranoyu plastidi roztashovuyetsya shar mureyinu rechovini harakternoyi dlya klitinnih obolonok bilshosti evbakterij i zokrema sino zelenih vodorostej Viddil glaukocistofitovih ye slipoyu gilkoyu evolyuciyi roslin dzherelo Plastidi chervonih vodorostej rodoplasti takozh zberigayut deyaki yaskravi oznaki sporidnenosti iz sino zelenimi vodorostyami cianobakteriyami zokrema tak zvani fikobilinovi pigmenti Plastidi zelenih vodorostej hloroplasti zberigayut najmenshe fenotipichnih ris shozhosti iz prokariotichnim endosimbiontom Vid zelenih vodorostej berut pochatok nazemni roslini prichomu vsi voni zberigayut pervinno simbiotichni hloroplasti Vtorinni endosimbiozi Chervoni ta zeleni vodorosti neodnorazovo stavali endosimbiontami geterotrofnih eukariotiv iz riznih carstv diskokristat tubulokristat ta platikristat Simbiozi sho vidbuvalisya za scenariyem eukariotichnij hazyayin eukariotichnij endosimbiont nazivayut vtorinnimi Vnaslidok vtorinnih endosimbioziv vinikla velika grupa viddiliv vodorostej iz vtorinno simbiotichnimi plastidami Vtorinno simbiotichni plastidi na vidminu vid pervinno simbiotichnih vkriti perevazhno chotirma membranami z yakih vnutrishnya membrana ye zalishkom plazmalemi sino zelenoyi vodorosti pervinnogo endosimbiontu druga membrana vidozminenoyu membranoyu travnoyi vakuoli gospodarya pervinnogo endosimbionta chervonoyi abo zelenoyi vodorosti yakij ye vodnochas vtorinnim endosimbiontom tretya membrana vlasne plazmalemoyu vtorinnogo endosimbionta chetverta zovnishnya membrana plazmalemoyu gospodarya vtorinnogo endosimbionta Citoplazma yaka perebuvaye u promizhku mizh drugoyu ta tretoyu membranoyu tak zvanij periplastidnij prostir ye redukovanoyu citoplazmoyu vtorinnogo endosimbionta i u deyakih vipadkah zberigaye 80S ribosomi ta redukovanu endoplazmatichnu sitku Zovnishni tretyu ta chetvertu membrani vtorinno simbiotichnih plastid nazivayut hloroplastnoyu endoplazmatichnoyu sitkoyu Prote osoblivo cikavim ye toj fakt sho u dvoh viddilah vodorostej iz vtorinno simbiotichnimi plastidami hlorarahniofitovih Chlorarachniophyta ta kriptofitovih Cryptophyta u periplastidnomu prostori zberigayetsya navit redukovane yadro vtorinnogo endosimbionta nukleomorf Nukleomorf otochenij dvomembrannoyu obolonkoyu i mistit DNK yaka za nayavnimi v nukleomorfi genami u hlorarahniofitovih sporidnena z yadernoyu DNK zelenih vodorostej a u kriptofitovih iz yadernoyu DNK chervonih vodorostej U deyakih vodorostej zokrema v evglenovih vtorinno simbiotichni plastidi vkriti ne chotirma a lishe troma membranami Vvazhayetsya sho u cih vodorostej odna z zovnishnih membran plastidi bula vtorinno vtrachena Vidomi vipadki koli plastidi yak pervinno tak i vtorinno simbiotichni redukuvalisya i todi vinikali vtorinno geterotrofni taksoni Zokrema dovedena vtorinna redukciya plastid u tipu Apicomplexa prashurami yakih buli fotoavtotrofni dinofitovi vodorosti Dinophyta Ye takozh dani yaki dozvolyayut pripustiti sho oomikotovi Oomycota utvorilis vnaslidok vtorinnogo perehodu do geterotrofnogo tipu zhivlennya vid odniyeyi z grup vodorostej tubulokristat Krim togo vtorinno geterotrofni taksoni nizkih ta serednih rangiv vid vidiv do poryadkiv i navit klasiv vidomi majzhe v kozhnomu viddili vodorostej a takozh sered vishih roslin dzherelo Poshirennya plastid cherez vtorinni endosimbiozi obumovilo viniknennya znachnoyi kilkosti viddiliv vodorostej Napriklad yaksho vodorosti z pervinno simbiotichnimi plastidami predstavleni lishe troma viddilami Glaucocystophyta Rhodophyta Chlorophyta to vodorosti zi vtorinno simbiotichnimi plastidami nalezhat do 12 viddiliv Sered nih dva viddili zberigayut nukleomorf Chlorarachniophyta Cryptophyta 8 viddiliv mayut chotirimembranni vtorinno simbiotichni plastidi bez nukleomorfu sho pohodyat vid chervonih vodorostej Raphidophyta Chrysophyta Eustigmatophyta Xanthophyta Phaeophyta Bacillariophyta Dictyochophyta Haptophyta Evglenofitovi vodorosti mayut trimembranni plastidi prashurami yakih ye zeleni vodorosti U Dinophyta plastidi utvoryuvalis neodnorazovo yak vnaslidok simbioziv iz zelenimi vodorostyami tak i navit vnaslidok tretinnih simbioziv iz vtorinno simbiotichnimi fotoavtotrofami RozmnozhennyaPodil klitini vidbuvayetsya bezstatevo mitozom proces dozvolyaye kozhnomu dochirnomu yadru otrimuvati po odnij kopiyi kozhnoyi hromosomi Cikl zhittya bagatoh eukariotiv vklyuchaye statevu reprodukciyu zminyuyuchis z gaployidnoyi fazi de v kozhnij klitini nayavna odna kopiya kozhnoyi hromosomi na diployidnu fazu de v kozhnij klitini nayavni po dvi kopiyi kozhnoyi hromosomi Diployidna faza yavlyaye soboyu zlittya dvoh gaployidnih gamet sho formuyut zigotu kotra v svoyu chergu mozhe dilitis v mitozi abo projti hromosomne skorochennya v mejozi U comu shabloni dopustimi znachni variaciyi u tvarin vidsutnya bagatoklitinna gaployidna faza prote kozhne pokolinnya roslin mozhe prohoditi cherez gaployidnu ta diployidnu bagatoklitinnu fazi V eukariotiv vidnoshennya ploshi do ob yemu menshe nizh u prokariotiv tomu metabolichnij proces ta proces utvorennya klitini trivaye dovshe Evolyuciya procesu statevoyi reprodukciyi ce pervisna ta fundamentalna harakteristika eukariotichnih organizmiv Bazuyuchis na filogentichnomu analizi molekulyarni biologi Daks ta Rodzher pripustili dzherelo sho stateve rozmnozhennya hoch i ne bulo osnovnim sposobom rozmnozhennya vse zh bulo pritamanne spilnomu predku vsih eukariotiv Osnovnij komplekt geniv kotri figuruyut na stadiyi mejozu nayavnij navit u takih parazitichnih vidiv yak Trichomonas vaginalis ta Giardia intestinalis kotri donedavna vvazhali aseksualnimi Oskilki ci dva vidi ye nashadkami rodiv sho na rannij stadiyi viddililis vid golovnogo evolyucijnogo dereva eukariotiv bulo pripusheno sho osnovni geni yaki figuruyut v fazi mejozu a otzhe i stateve rozmnozhennya buli shvidshe za vse pritamannimi spilnomu predku eukariotiv Eukariotichni vidi sho pomilkovo vvazhalisya bezstatevimi napriklad parazit z rodu Leishmania vse zh mayut statevij cikl Takozh pevni chinniki vkazuyut na te sho ameba kotra vvazhalas bezstatevim vidom z davnih chasiv rozmnozhuvalasya statevim shlyahom ta sho v bilshosti suchasnih bezstatevih vidiv neshodavno ta nezalezhno odin vid odnogo prokinuvsya seksualnij potyag dzherelo sumnivno obgovoriti Dzherela vidpov redaktori D M Grodzinskij Kiyiv KMM 2012 744 s ISBN 978 966 1673 12 9 Archibald J M Cavalier Smith T Maier U amp Douglas S 2001 Molecular chaperones encoded by a reduced nucleus the cryptomonad nucleomorph J Mol Evol 52 490501 Kostikov I Yu Dzhagan V V Demchenko E M Bojko O A Bojko V R Romanenko P O Botanika Vodorosti ta gribi Kiyiv 2004 Thomas P Curtis William T Sloan and Jack W Scannel 6 PNAS 99 16 10494 10499 Arhiv originalu za 24 veresnya 2015 Procitovano 19 serpnya 2008 Retallack G J Krull E S Thackray G D and Parkinson D 2013 Problematic urn shaped fossils from a Paleoproterozoic 2 2 Ga paleosol in South Africa Precambrian Research 235 71 87 doi 10 1016 j precamres 2013 05 015 Sergej Yastrebov 22 11 2017 Novosti nauki na Elementah Arhiv originalu za 12 travnya 2020 Procitovano 12 lipnya 2020 ros P J Keeling W F Doolittle Alpha tubulin from early diverging eukaryotic lineages and the evolution of the tubulin family Molecular Biology and Evolution 13 1297 1305 doi 10 1093 oxfordjournals molbev a025576 PMID 8952074 Vossbrinck C R J V Maddox S Friedman B A Debrunner Vossbrinck and C R Woese 1987 Ribosomal RNA sequence suggests microsporidia are extremely ancient eukaryotes Nature 326 411 414 doi 10 1038 326411a0 PMID 3550472 Posilannyahttp mbe oxfordjournals org cgi reprint 13 10 1297 http itol embl de itol cgi 8 lipnya 2007 u Wayback Machine http www ucmp berkeley edu alllife eukaryota gif 13 lipnya 2007 u Wayback Machine EUKARIOTI 12 bereznya 2016 u Wayback Machine Farmacevtichna enciklopediya Ce nezavershena stattya z biologiyi Vi mozhete dopomogti proyektu vipravivshi abo dopisavshi yiyi